

人体における代謝過程では酸,およびより少量の塩基が持続的に産生されている。水素イオン(H+)は特に反応性が高い;これは負電荷をもつタンパク質に接着し,高濃度ではタンパク質全体の電荷,構造,機能を変えてしまうこともある。細胞の機能を維持するために,生体は血中H+濃度を狭い範囲内に維持する精巧な機序を備えており,その範囲は典型的には37~43nEq/L(37~43nmol/L),すなわちpH 7.37~7.43(ここでpH = −log[H+])である。H+濃度が40nEq/L(40nmol/L),すなわちpH = 7.40の状態が理想である。この機序が障害されると臨床的に重大な影響が生じる可能性がある。
酸塩基平衡は水分の代謝および電解質バランスと密接に結びついており,これらのシステムのうち1つが障害されるとしばしば他のシステムにも影響が生じる。
酸塩基の生理
大半の酸は以下の代謝に由来する:
炭水化物
脂肪
炭水化物および脂肪の代謝により,1日当たり15,000~20,000mmolの二酸化炭素(CO2)が発生する。CO2はそれ自体は酸ではないが,炭酸脱水酵素に属する酵素の存在下で,CO2は血中の水(H2O)と結合して炭酸(H2CO3)を産生し,これが水素イオン(H+)と重炭酸イオン(HCO3−)に解離する。H+は赤血球中でヘモグロビンに結合し,肺胞における酸素化に伴って放出されるが,このときに別の炭酸脱水酵素により反応が逆向きに進み,水(H2O)およびCO2が産生されて,前者は腎臓から排出され,後者は呼吸のたびに吐き出される。
比較的少量の有機酸が以下に由来して生じる:
グルコースおよび脂肪酸の不完全代謝による乳酸およびケト酸
硫黄含有アミノ酸(システイン,メチオニン)の代謝による硫酸
陽性荷電アミノ酸(アルギニン,リジン)の代謝
食物中のリン酸の加水分解
この「固定」または「代謝による」酸は呼出されないため,中和または腎から排出されなければならない。
大半の塩基は以下に由来する:
陰性荷電アミノ酸(グルタミン酸およびアスパラギン酸)の代謝
乳酸,クエン酸などの有機陰イオンの酸化および消費(これらによりHCO3−が産生される。)
酸塩基平衡
酸塩基平衡は以下によって維持される:
化学的緩衝作用
肺の活動
腎臓の活動
化学的緩衝作用
化学的緩衝物質はpHの変化に抵抗する溶液である。細胞内および細胞外の緩衝物質は酸塩基平衡異常に即座に反応する。骨も,(特に酸負荷時に)重要な緩衝作用を有する。
緩衝物質は弱酸とその共役塩基から成る。共役塩基はH+を受容でき,弱酸はそれを放出できるため,それにより遊離H+濃度の変化が最小限に抑えられる。緩衝系は,その平衡定数(pKa)付近で最も効果的にpHの変化を最小限に抑える;そのため,生体内には多数の緩衝系が存在していると思われるものの,生理的に重要であるのはその一部のみである。
緩衝系のpHと構成要素の濃度との関係は,以下のHenderson-Hasselbalchの式で表される:

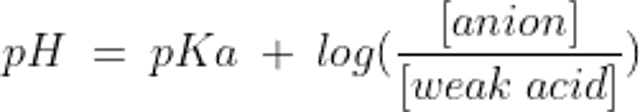
ここでpKaは弱酸の解離定数である。
細胞外の最も重要な緩衝物質はHCO3−/CO2系であり,以下の式で表される:

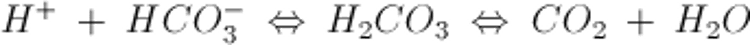
H+の増加は右向きの反応を促進してCO2を発生させる。
この重要な緩衝系は高度の調節を受けている;CO2濃度は肺胞換気によって,H+およびHCO3−の濃度は腎排泄によって微細に調節できる。
したがって,Henderson-Hasselbalchの式に当てはめると,この系におけるpH,HCO3−およびCO2の間の関係は以下のようになる:

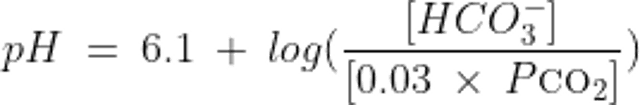
あるいは,Henderson-Hasselbalchの式から導かれるKassirer-Bleichの式では以下のように表される:

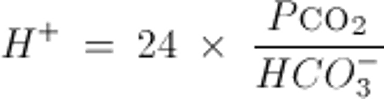
注:動脈血pHを[H+]に換算するには以下を用いる:


または

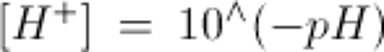
上記いずれの式も,酸塩基平衡は二酸化炭素分圧(Pco2)とHCO3−の比に左右され,いずれか単独の絶対値には影響されないことを表している。これらの式を用いると,いずれか2つの変数がわかれば第3の変数を算出することができる。
その他の重要な化学的緩衝物質としては,細胞内の有機および無機リン酸や,赤血球中のヘモグロビンを含むタンパク質などがある。重要性が比較的低いものに,細胞外リン酸および血漿タンパク質がある。
骨は酸負荷の消費後に重要な緩衝物質となる。骨は初めのうちは,H+との交換で炭酸水素ナトリウム(NaHCO3)および炭酸水素カリウム(KHCO3)を放出している。酸負荷が遷延すると,骨は炭酸カルシウム(CaCO3)およびリン酸カルシウム(CaPO4)を放出する。したがって,長期のアシデミアは骨の脱灰および骨粗鬆症の一因となる。

肺でのpH調節
CO2濃度は1回換気量および呼吸数(分時換気量)の変化により細かく調節されている。pHが低下すると,動脈化学受容器に感知され,それにより1回換気量または呼吸数が増加し,結果としてCO2が呼出され,血中pHが上昇する。化学的な干渉が直ちに起こるのとは対照的に,肺による調節には数分から数時間を要する。肺による調節は約50~75%の効果があるが,pHを完全に正常化することはできない。
腎臓でのpH調節
腎臓はHCO3−の排泄量または再吸収量を調整することでpHを調節している。HCO3−の再吸収は遊離H+の排出と等価である。腎臓での酸塩基処理の変化は,酸塩基平衡の状態が変化してから数時間から数日後に生じる。
血清中のHCO3−は,糸球体を通過する過程で全て濾過される。HCO3−は主に近位尿細管で再吸収されるほか,より程度は低いが集合管でも再吸収される。遠位尿細管細胞内のH2OはH+と水酸化物イオン(OH−)に解離するが,炭酸脱水酵素の存在下では,OH−はCO2と結合してHCO3−となり,これは傍尿細管毛細血管に送り返される一方,H+は尿細管腔内に分泌され,これが自由に濾過されたHCO3−と結合してCO2とH2Oになり,これらも再吸収される。したがって,遠位側で再吸収されたHCO3−イオンは新たに発生したものであり,濾過されたものと同じではない。
有効循環血液量が減少すると(利尿療法に伴って生じるものなど)HCO3−の再吸収が増加する一方,酸負荷に反応して副甲状腺ホルモンが増加するとHCO3−の再吸収は減少する。また,Pco2が上昇するとHCO3−再吸収の増加を招くのに対して,塩化物イオン(Cl−)が欠乏(通常体液量減少に起因する)すると,近位尿細管でのナトリウムイオン(Na+)の再吸収とHCO3−の産生が増加する。
酸は近位尿細管内および遠位尿細管内に能動的に排泄されて,そこで尿中緩衝物質―主として自由に濾過されたリン酸(HPO4−2),クレアチニン,尿酸,およびアンモニア―と結合して体外に輸送される。アンモニア以外の緩衝物質は決まった濃度で濾過され大量の酸負荷があると欠乏する可能性があるため,アンモニア緩衝系は特に重要である;対照的に,アンモニアの産生は,酸負荷の変化に応じて尿細管細胞により活発に調整される。動脈血pHは酸分泌の主な決定因子であるが,排泄はカリウム(K+),Cl−,およびアルドステロン濃度の影響も受ける。細胞内K+濃度とH+分泌とは相互に関連している;K+が欠乏するとH+分泌が増加し,その結果代謝性アルカローシスが生じる。






